

軟骨無形成症においてFGFR3シグナルの過剰活性化がCREBシグナルを介して静止層軟骨細胞のターンオーバーを障害する
| 著者: | Nanao Horike, Seiya Oura, Saeko Koyamatsu, Noriko Tanaka, Yuki Iimori, Kaori Fujita, Takahiro Nemoto, Masahito Ikawa, and Noriyuki Tsumaki |
|---|---|
| 雑誌: | Nature Communications, 2026年2月26日(オンライン公開), Volume 17, Article number: 1856 (2026), DOI:https://doi.org/10.1038/s41467-026-69507-9 |
- 軟骨無形成症
- FGFR3
- CREB
 研究室送別会(大阪府高槻市)後列中央(責任著者 妻木教授)、前列左端(小屋松 研究員)、前列左から4番目(松田 技術職員)、前列左から5番目(加藤 技術職員)、前列右から4番目(田中 研究員)、前列右端(筆頭著者 堀家)
研究室送別会(大阪府高槻市)後列中央(責任著者 妻木教授)、前列左端(小屋松 研究員)、前列左から4番目(松田 技術職員)、前列左から5番目(加藤 技術職員)、前列右から4番目(田中 研究員)、前列右端(筆頭著者 堀家)
 左:堀家、右:妻木教授
左:堀家、右:妻木教授
論文サマリー
骨の成長は、成長軟骨板と呼ばれる骨端部の軟骨組織で起こる。この成長軟骨板は、静止層・増殖層・肥大層の3層から構成されており、静止層の軟骨細胞が分化して増殖層軟骨細胞を供給し、その後に骨組織へと置き換わることで骨が縦方向に伸びる仕組みとなっている。
軟骨無形成症は、FGFR3(線維芽細胞増殖因子受容体3型)遺伝子の活性型変異によって引き起こされる疾患であり、四肢短縮型低身長を主な症状とする。FGFR3は軟骨細胞に高発現し、細胞の増殖と分化を調節する受容体型チロシンキナーゼである。軟骨無形成症では、この受容体が過剰に活性化することで成長軟骨板の機能が障害されるが、各分化段階の軟骨細胞における詳細なメカニズム、特に静止層軟骨細胞から増殖層軟骨細胞への供給を制御する仕組みはこれまで明らかになっていなかった。
本研究では、軟骨無形成症の原因変異(p.Gly380Arg)をノックインしたモデルマウス(Fgfr3Achマウス)を作製した。このモデルマウスは全身の骨長短縮を呈し、軟骨無形成症の病態を再現した。詳細な解析により、これまで報告されていた異常に加え、成長軟骨板の静止層が拡大していることを新たに発見した。EdUラベリングおよび細胞系譜追跡解析を行ったところ、FGFR3の過剰活性化によって静止層軟骨細胞のターンオーバーが障害され、細胞分裂後に静止層にとどまり続けることで増殖層への供給が阻害されることが明らかとなった。
さらに、成長軟骨板のシングルセルRNAシーケンス解析と免疫組織化学解析を組み合わせることにより、拡大した静止層に対応する細胞クラスターを同定した。パスウェイ解析および機能実験から、FGFR3の下流でCREBが活性化されることが静止層軟骨細胞の幹細胞様特性を損なうことで骨成長障害に寄与していることを解明した。実際に、CREB阻害剤(666-15)をモデルマウスに投与したところ、成長軟骨板の病理所見が改善し、骨長短縮の表現型が回復した。
本研究の成果により、軟骨細胞分化を制御する新たな分子機構として「FGFR3/CREB経路による静止層軟骨細胞のターンオーバー制御」が明らかになった。静止層軟骨細胞の振る舞いが骨成長に果たす役割の理解が分子レベルで深まるとともに、CREBが軟骨無形成症の新たな創薬標的分子となる可能性が示された。今後、この知見が軟骨無形成症をはじめとする難治性軟骨疾患に対する新規治療法の開発に貢献することが期待される。
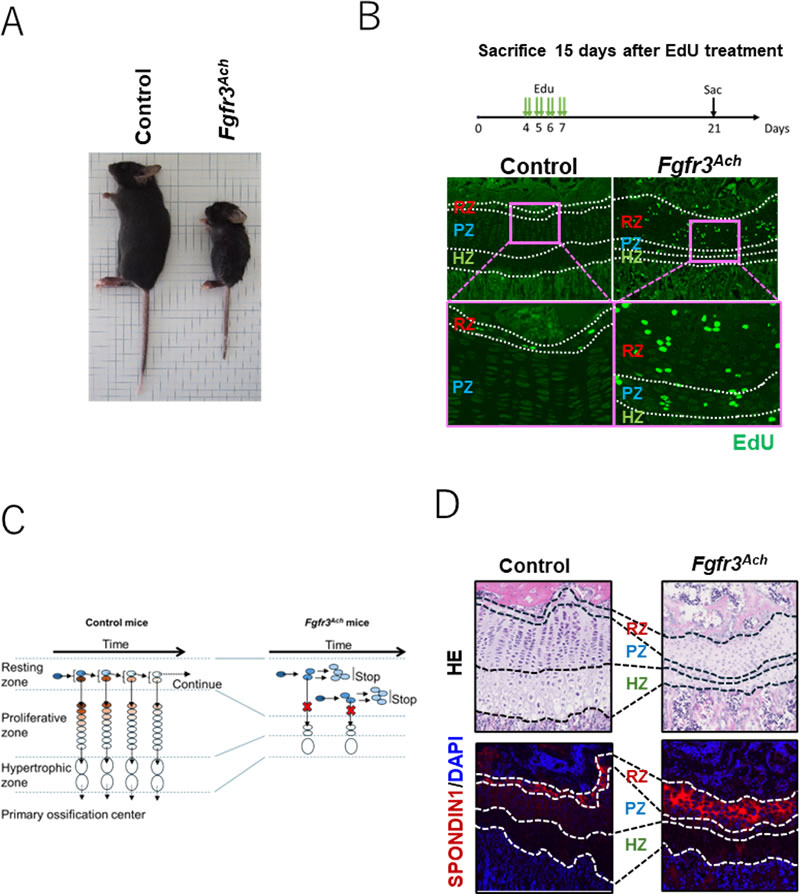
A:Fgfr3 Ach マウスは対照マウスより全身の骨長が短く、軟骨無形成症の低身長表現型を再現した。
B:生後4~7日目のマウスにEdUを投与しP21まで追跡した。EdUは細胞分裂時にDNAに取り込まれるため、分裂後も長期間細胞内にEdUを保持している細胞をLRCs(Label-retaining chondrocytes:標識保持軟骨細胞)と呼び、ターンオーバーの遅い静止層軟骨細胞のマーカーとなる。対照マウスでは静止層のLRCsはごく少数だったが、Fgfr3 Achマウスでは拡大した静止層に多数のLRCsが認められ、静止層軟骨細胞のターンオーバー障害と増殖層への供給阻害が示された。
C:対照マウスでは静止層軟骨細胞が幹細胞様に分裂し増殖層へ供給された。Fgfr3 Ach マウスでは娘細胞が静止層に蓄積し増殖層・肥大層が減少、骨成長が妨げられた。
D:FGFR3下流のCREBシグナルで発現制御されるSPONDIN1が、Fgfr3 Achマウスの拡大した静止帯で特異的に強く発現した。
著者コメント
本研究では、軟骨無形成症モデルマウスの成長軟骨板をシングルセルRNAシーケンス解析することにより、FGFR3がCREBを介して静止層軟骨細胞のターンオーバーを制御するという新たなメカニズムを解明することができました。CREBは副作用の懸念から直接の治療標的としては難しい面もありますが、今回明らかにした経路をさらに掘り下げることで、副作用の少ない新たな分子標的の同定につなげ、軟骨無形成症の根本的な治療薬開発の発展に貢献できればと考えています。最後に、着手から結果をまとめるまでに長い時間を要しましたが、丁寧にご指導くださいました妻木範行教授、ならびに共著者の先生方・研究室の皆様に、この場を借りて心より感謝申し上げます。
堀家 なな緒 (大阪大学大学院 生命機能研究科 個体機能学講座 組織生化学研究室/医学系研究科 生化学・分子生物学講座(組織生化学))
2026年6月30日